

LUCA – Last universal common ancestor – mit Gegenwartsbezug
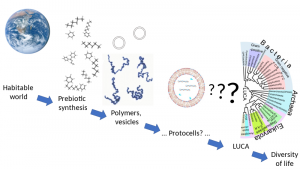
Die Entstehungsstadien des Lebens reichen von gut verstandenen Stadien wie der bewohnbaren Erde und der abiotischen Synthese einfacher Moleküle bis hin zu weitgehend unbekannten Stadien wie der Ableitung des letzten universellen gemeinsamen Vorfahren (LUCA) mit seinen komplexen molekularen Funktionalitäten.
Es gab eine Zeit, in der es kein Leben auf der Erde gab, und es gab eine Zeit, in der es DNA-vererbende Zellen gab.
„Deshalb sollte ich aus der Analogie schließen, dass wahrscheinlich alle organischen Wesen, die jemals auf dieser Erde gelebt haben, von einer Urform abstammen, der zuerst Leben eingehaucht wurde.“ Charles Darwin, On the Origin of Species, 1859
Seit der Veröffentlichung von Darwins Hauptwerk, „The Origin of Species“, rätseln Wissenschaftler über die Natur seiner angedeuteten Urgestalt (primordial form).
Unterdessen trägt die Lösung den Namen LUCA , ein Akronym für Last universal common ancestor, also letzter universeller gemeinsamer Vorfahre.
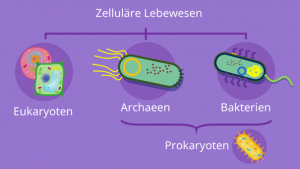
Abb. Eukaryoten sind nicht nur große, komplexe Archaeen; Genomisch und auf zellulärer Ebene sind sie echte Chimären, da sie archaeische Ribosomen im Zytosol und bakterielle Ribosomen in den Mitochondrien besitzen.
LEBEN IST EINE OPTION
Die Erde ist in der Anfangszeit des Sonnensystems vor 4,5 Milliarden in einem regelrechten Gesteinsbombardement aus dem All, als ein mondgroßer Gesteinsbrocken auf die Erde einschlug und die Erde in eine Kugel aus siedender Lava verwandelte. Magma-Ozeane mit Temperaturen über 2.000 °K wandelten das gesamte Wasser aus der frühen Ansammlung in die Gasphase in atmosphärisches Kohlendioxid (CO 2) um.
So musste die Kohlenstofffixierung genutzt werden, bei der anorganischer Kohlenstoff in organische Kohlenstoffverbindungen umgewandelt werden, die vom Leben genutzt werden können. Vereinfacht ausgedrückt wird die Kohlenstofffixierung von Bakterien und Archaeen übernommen und reduziert Wasserstoff und Kohlendioxid zu Kohlenmonoxid und Ameisensäure, die dem Leben nützlich sind.
So entstanden irgendwo auf der von Ozeanen bedeckten frühen Erde die ersten Zellen.
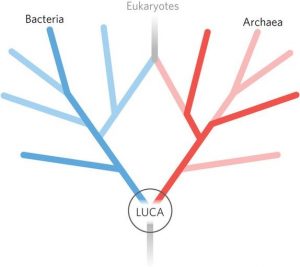
Weil der genetische Code und die Aminosäurechiralität universell sind, gehen alle modernen Lebensformen letztlich auf diese Evolutionsphase zurück. Das war die Zeit, in der der letzte universelle gemeinsame Vorfahre (LUCA) aller Zellen lebte.
DAS GRÖSSTE BIOLOGISCHE EREIGNIS FAND VOR MEHR ALS 3,5 MILLIARDEN JAHREN STATT- es ist die Entstehung des Lebens
Es muss beachtet werden, dass LUCA nicht der Ursprung des Lebens ist. Der früheste Beweis für Leben stammt aus der Zeit vor 3,7 Milliarden Jahren in Form von Stromatolithen, bei denen es sich um Sedimentschichten handelt, die von Mikroben abgelagert wurden. Vermutlich gab es schon vorher Leben. Doch die Ankunft von LUCA und seine Entwicklung zu Archaeen und Bakterien könnten zu jedem Zeitpunkt vor 2 bis 4 Milliarden Jahren stattgefunden haben.
In den allerersten Phasen des Lebens auf der Erde entstanden Leben, Genetik und Epigenetik aus den Elementen.
Aus damals wie heute hochverfügbarem Basalt und einfachen Wärmeflüssen konnte das für die RNA-Faltung richtige Verhältnis zwischen Magnesium- und Natriumionen durch natürliche Prozesse bereitgestellt werden. In den feinen Kanälen von basaltischem Glas reicherten sich Magnesiumionen stark an. Thermophoretische Bedingungen verbesserten die Selbstreproduktion eines Ribozyms, – Ribozyme sind katalytisch aktive RNA-Moleküle, die wie Enzyme chemische Reaktionen katalysieren und unter dem Einfluss der Thermophorese in der Lage sind, mehrere kurze RNA-Stränge zu verknüpfen und damit sehr lange RNA-Moleküle zu erzeugen. Diese RNA-Stränge als Ribozyme brauchten, wie eingangs erwähnt Magnesiumionen für ihre katalytischen Aktivitäten, vertrugen aber keine hohen Konzentrationen an Natriumionen.
LUCA’s Stoffwechsel war: anaerob, CO2‐ und N2‐fixierend, H2‐abhängig und thermophil
Sein Stoffwechsel ist voll von O 2 -empfindlichen Enzymen. Dazu gehören Proteine, die reich an O 2 -empfindlichen Eisen-Schwefel-Clustern (FeS) sind, und Enzyme, deren Reaktionsmechanismen die Erzeugung von Radikalen (ungepaarten Elektronen) über S-Adenosylmethionin (Ademetionin) beinhalten. Das passt gut zu der 50 Jahre alten, aber immer noch modernen Ansicht, dass FeS-Cluster sehr alte Cofaktoren im Stoffwechsel darstellen. Es passt auch zu neueren Erkenntnissen über die alte und spontane (nichtenzymatische) Chemie, die der S-Adenosylmethionin- (Ademetionin) -Synthese zugrunde liegt.
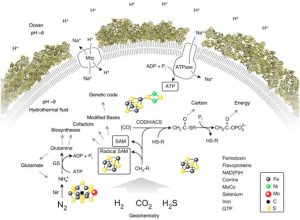
Abb. Rekonstruktion des letzten gemeinsamen Vorfahren aus Genomsequenzen. Übergangsmetallcluster sind symbolisiert. Methylgruppen, CH3-R; CODH/ACS, Kohlenmonoxid Dehydrogenase/Acetyl-CoA-Synthase; GS, Glutaminsynthetase; HS-R, organische Thiole; LUCA, letzter universeller gemeinsamer Vorfahre; Mrp, Na+ /H+ -Antiporter vom MrP-Typ; Nif, Nitrogenase; SAM, S-Adenosylmethionin (Ademetionin)
Quelle: Neuauflage von Weiss et al. (2018) unter der Creative Commons Attribution Licence. Einzelheiten zur Physiologie von LUCA siehe Weiss et al. (2016) und Weiss et al. (2018).
355 Gene verbinden LUCA als gemeinsamer Vorfahre mit Bakterien und Archaeen
Die 355 Gene deuteten ziemlich genau auf einen Organismus hin, der unter den Bedingungen lebte, die in Tiefseequellen herrschen, den gashaltigen, metallbeladenen, äußerst heißen Quellen, die durch die Wechselwirkung von Meerwasser mit durch den Meeresboden ausbrechendem Magma entstehen.
Das Leben ist eine chemische Reaktion. Energieerhaltung (ATP-Synthese) und Energienutzung (ATP-Hydrolyse). Es ist die Energiegewinnung, die die Zellen lebendig macht.
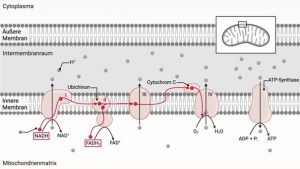
Der Energiestoffwechsel ist wichtig, da die Aufnahme von Nährstoffen und deren Umwandlung in die Bestandteile des Lebens eine Energiezufuhr erfordern. Ohne Energiefluss ist die Zelle nur eine Ansammlung von Molekülen, die zum endgültigen Zerfall verurteilt ist. Alle Zellaktivitäten – metabolische, sensorische oder andere – erfordern eine Energieumwandlung und die Zufuhr von Energie in einer Form, die von anderen Prozessen in der Zelle genutzt werden kann.
Alles Leben, das wir kennen, verwendet Phosphat, hauptsächlich als ATP, aber auch als GTP, Phosphoenolpyruvat und Acylphosphat. Dies bedeutet normalerweise die Hydrolyse einer Phosphoanhydridbindung in ATP zu ADP und Phosphat (Pi) oder wie in manchen Reaktionen zu AMP und PPi. Das an einem Tag über die Atmungskette produzierte ATP entspricht dem eigenen Körpergewicht. Der ATP-Aufwand bei der Phosphoanhydridhydrolyse bedeutet, dass die Anhydridbindung in ATP resynthetisiert werden muss, was die Aufgabe des Energiestoffwechsels ist.
Ganz allgemein ist die Energiegewinnung die wichtigste Reaktion in Zellen, da ohne Energieerhaltung alles andere unerheblich ist und mitochondriale Dysfunktion zur Folge hat.
Neben der grundlegenden Rolle der Mitochondrien bei der Energieerzeugung (ATP) sind die Mitochondrien auch die Hauptproduzenten von freien Sauerstoffradikalen (ROS). Wenn die Abwehrmechanismen der Antioxidantien geschwächt sind, lösen diese reaktiven oxidativen Spezies (ROS) eine Kaskade schädlicher Ereignisse im Mitochondrium aus.
Neben ATP gibt es noch andere Energiewährungen, aber ATP ist bei weitem die wichtigste.
Die Theorie der Eisen-Schwefel-Welt, die die Umwandlung von FeS in FeS 2 als eine wichtige energieliefernde Reaktion für die Entstehung von Leben postuliert, dass die exergonische Reaktion von FeS mit H 2 S zur Bildung von FeS 2 (Pyrit) und H 2 eine frühe Form des Energiestoffwechsels auf der Urerde war. Seit dem Archaikum spielt die sedimentäre Pyritbildung eine wichtige Rolle in den globalen Eisen- und Schwefelkreisläufen mit direktem Einfluss auf die Redoxchemie der Atmosphäre.
Seit 3.5-3.8 Mrd. Jahre ist der Methylgruppenspender für die RNA, DNA und Histone, das S-Adenosyl-L-Methionin aus den Genomsequenzen von LUCA bekannt
Neben Cobalamin-abhängigen Methyltransferasen wird die größte Familie innerhalb der Transferasen durch S-Adenosyl-L-Methionin (Ademetionin) abhängige Methyltransferasen (MTs) gebildet. Diese MTs sind in der Natur allgegenwärtig und methylieren unter S-Adenosyl-L-Methionin (Ademetionin) -Verbrauch und der Bildung von S-Adenosyl-L-Homocystein als Nebenprodukt eine Vielzahl von Substraten. Das Substratspektrum von MTs beinhaltet unter anderem DNA, RNA, Proteine, Peptide und kleine Moleküle: DNA- und RNA-Methyltransferasen methylieren Cytosin bzw. Adenin. Protein-Methyltransferasen methylieren Lysin und Arginin. C-, O- und N-Methyltransferasen für kleine Moleküle methylieren u.a. Steroide, Geschmacks- und Geruchsstoffe, sowie Alkaloide.
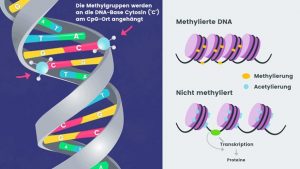
Die DNA-Methylierung spielt eine wichtige Rolle bei der epigenetischen Signalübertragung und hat Auswirkungen auf die Genregulation, die Chromatinstruktur, die Entwicklung und von Krankheiten. DNA-MTs wirken regulatorisch auf die Epigenetik von Organismen ein,
Methyltransfer von S-Adenosylmethionin (Ademetionin)
Der Methyltransfer von S-Adenosylmethionin (Ademetionin) ist vielleicht seine bekannteste Rolle, da er am Zwischenstoffwechsel und an der Modifikation von Nukleinsäuren und Proteinen beteiligt ist. Die DNA-Methylierung bildet eine Grundlage für das aufstrebende Gebiet der Epigenetik, während eine fehlerhafte DNA-Methylierung bei Erkrankungen häufig vorkommt, wobei wie bei den Krebserkrankungen Fehler mit Veränderungen in der DNA-Replikation und -Transkription verbunden sind.
S- Adenosyl-L-Methionin (Ademetionin) ist ein wichtiger biologischer Methyldonor [Cantoni GL . S-Adenosylmethionin – ein neues Zwischenprodukt, das enzymatisch aus L-Methionin und Adenosintriphosphat gebildet wird . J Biol Chem . 1953 ; 204 : 403 – 16 .] und spielt eine Schlüsselrolle bei der Methylierung von DNA, RNA, Histonen, Proteinen und einer Vielzahl kleiner Moleküle [ [Markham GD . S-Adenosylmethionin in der Nature Encyclopedia of Life Sciences . London : Nature Publishing Group; 2002 .].
In einer anderen Familie von Signalwegen führt die Decarboxylierung von S-Adenosylmethionin (Ademetionin), gefolgt von der Übertragung der Propylamin-Einheit, zu den Polyaminen Spermin und Spermidin, die bei der Regulierung der Zellproliferation verwendet werden. Die Übertragung einer Methylgruppe von S -Adenosylmethionin ergibt S -Adenosylhomocystein, das mehrere Methyltransferasen wirksam hemmt.
Der Transsulfurierungsweg dient u.a. der Glutathionsynthese .
S-Adenosylmethionin (Ademetionin) und seine Metaboliten spielen eine Vielzahl von Rollen im Zellleben. S-Adenosylmethionin (Ademetionin) ist eines der wenigen Sulfoniumionen, die in der Natur vorkommen, und das kationische Zentrum verleiht ihm eine chemische Vielseitigkeit, die nur wenige andere biologische Einheiten erreichen, sodass es als Alkylierungsmittel und Vorläufer freier Radikale sowie als Regulierungsmittel fungieren kann.
Die Idee, dass Radikale eine wesentliche Rolle in der Biologie spielen könnten, war viele Jahre lang umstritten. In den 1980er Jahren erkannte man, dass organische Radikale zwar hochreaktiv und im „freien“ Zustand potenziell gefährlich sind, aber über eine einzigartige und starke Reaktivität verfügen, die von der Biologie ausgenutzt werden kann, wenn die Radikale angemessen eingeschränkt werden können. Seitdem sind Radikale als wichtige Akteure in biochemischen Reaktionen anerkannt, sowohl als Schlüsselreaktionszwischenprodukte als auch als katalytisch essentielle Protein-Cofaktoren.
Zu den frühesten Cofaktoren, die an biologischen Radikalreaktionen beteiligt waren, gehörte Adenosylcobalamin (Coenzym B 12).
S -Adenosyl-L-Methionin ist im Vergleich zu Adenosylcobalamin (Coenzym B 12) ein viel einfacherer, rein organischer Cofaktor und verfügt über eine 5′-Desoxyadenosyl-C-S-Bindung zum Sulfoniumschwefel von S -Adenosyl-L-Methionin anstelle der kritischen 5′-C-Co-organometallischen Bindung, die die Adenosyleinheit zu Cobalamin verbindet.
Radikal-SAM (Radical-Adenosylmethionin) ist eine Bezeichnung für eine Superfamilie von Enzymen, die einen [4Fe-4S] +- Cluster verwenden, um S -Adenosyl- L -methionin reduktiv zu spalten, um ein Radikal zu erzeugen , normalerweise ein 5′- Desoxyadenosylradikal (5′-dAdo). ), als kritisches Zwischenprodukt.
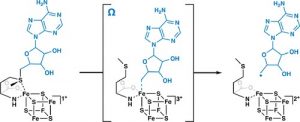
Radikale SAM-Enzyme sind an der Cofaktor- Biosynthese, Enzymaktivierung, Peptidmodifikation sowie posttranskriptionellen und posttranslationalen Modifikationen beteiligt, Bildung von Metalloprotein- Clustern, tRNA- Modifikation, Lipidstoffwechsel, Biosynthese von Antibiotika und Naturstoffen usw. Die überwiegende Mehrheit der bekannten radikalischen S -Adenosyl-L-Methionin-Enzyme gehört zur radikalen SAM-Superfamilie.
Nützliche Freundschaften von Bakterien und Eukaryont
ATP-Produktion durch Chloroblasten und Mitochondrien
Wir alle haben den gleichen Ursprung und sind mit sämtlichen Lebewesen dieser Erde aufgrund unserer Gene verwandt. Wir befinden uns in gegenseitiger Abhängigkeit mit allen Lebewesen sowie den geologischen und atmosphärischen Gegebenheiten.
Etwa vier Milliarden Jahre brauchte es, bis auf der Erde Tiere, Menschen und Pflanzen entstehen konnten. Zwei große biologische Revolutionen waren dazu notwendig:
Vor mehr als 2,5 Milliarden Jahren vollzieht das Leben auf der Erde einen entscheidenden Schritt: Bisher bezogen die Einzeller ihre Lebensenergie aus einer Vielzahl chemischer, lichtunabhängiger Prozesse, wie auch heute noch. Nun entwickeln sie zusätzlich die Photosynthese, durch die sie, wie später die Pflanzen, mithilfe der Lichtenergie aus Kohlendioxid und Wasser Zucker bilden. So wird das in Chloroblasten durch Photosynthese erzeugte ATP die primäre Quelle für biologisch nützbare Energie auf unserer Erde.
Der zweite alles entscheidende Schritt war der Transfer dieser neuen Erfindung der ATP-Erzeugung durch Endobiose, einer Gemeinschaft aus Organismen unterschiedlicher Art, bei der ein Organismus in einem anderen lebt (Endosymbiose), ohne diesen zu schädigen. Diese Zufallsinnovation fand vor 1,5 Milliarden Jahren statt. Die Natur hatte das Mitochondrium erfunden.
Eukaryoten sind aus einer Symbiose hervorgegangen, bei der Archaeen als Wirtszellen Bakterien aufgenommen haben, die sich dann zu Mitochondrien entwickelten, den Energielieferanten der eukaryotischen Zellen. Eine Hypothese besagt, dass das Wirts-Archaeon für seinen Stoffwechsel Wasserstoff benötigte, und dass die Vorläufer der Mitochondrien Wasserstoff produzierten.
Nach dieser Endosymbiontentheorie entstanden unsere Lebensenergieproduzenten, die Mitochondrien vor 1,5 Milliarden Jahren durch Endobiose aus einem Alpha-Proteobakterium und der Archaeen als Wirtszellen, die das Bakterium inkorperierten.
Die entscheidende Funktion des Mitochondriums ist die Herstellung von Energie in Form von Adenosintriphosphat (ATP) aus Wasserstoff und Sauerstoff, die die Anpassung an die zunehmend sauerstoffreiche Atmosphäre ermöglichte.
Dies war auch die Schlüsselinnovation auf dem Weg zum multizellulären Leben. Es gab nun genügend Energie, um sich den Luxus von Organen mit hohem Energieverbrauch leisten zu können. Die ausdifferenzierte und physiologischen Funktionen unserer Organe: Gehirn, Herz, Skelettmuskel, Retina, Leber u.a. wären ohne mitochondrialer Energiebereitstellung (Atmungskette) undenkbar. Ohne Mitochondrien wären wir in großen Schwierigkeiten, denn es gäbe eine individuelle Energiekrise, die mit dem strukturierten und ausdifferenzierten Leben nicht vereinbar wäre, wie auch eine globale biologische Energiekrise, wenn es keine Chloroplasten der Pflanzen gäbe.
Eine Besonderheit der Mitochondrien ist ihre duale genetische Kontrolle durch mitochondriale DNA (mtDNA) – DNA der inkorperierten Bakterien-und nukleare DNA (nDNA)-DNA der Archaeen als Wirtszelle.
99% aller strukturellen und funktionellen Proteine des Mitochondriums sowie die meisten für Transkription, Translation und Replikation des mitochondrialen Genoms erforderlichen Proteine werden durch die nukleare DNA (nDNA) kodiert. Nur 1% der Proteine sind in der mitochondrialen
DNA (mtDNA), also außerhalb des Zellkerns, im Mitochondrium selbst verschlüsselt.
Die mitochondriale DNA (mtDNA) ist als doppelsträngige und zumeist als zirkuläre DNA in der Matrix der Mitochondrien zu finden. Sie wurde 1964 von Ellen Haslbrunner, Hans Tuppy und Gottfried Schatz, – am Institut für Biochemie der medizinischen Fakultät der Universität Wien, aufgrund biochemischer Messungen entdeckt.
S-Adenosyl-L-Methionin (Ademetionin) kann auch mit Hilfe des S-adenosylmethionine mitochondrial carrier protein (SAMC) in die Mitochondrien gelangen, wo es von DNA-Methyltransferasen (DNMTs) zur Methylierung der mitochondriale DNA (mtDNA) verwendet wird.
S-Adenosyl-L-Methionin als Methylgruppen-Spender an die Erbsunstanz
ATP war ausreichend bei der Biosynthese von S-Adenosyl-L-Methionin, dem „energetisierten Methionin“, vorhanden, dass auch der epigenetische Marker CH3- zur Trans-Methylierung der Erbsubstanz bereitstand. Die Epigenetik begann die Genetik zu ergänzen und das Leben zu bereichern.
Das Mikrobiom
Die Mikroorganismen, aus denen unser Mikrobiom besteht, kommen hauptsächlich in sieben Körperregionen vor: Magen, Nase, Mund, Lunge, Haut, Dickdarm und Geschlechtsorgane – und jeder Mensch besitzt in jeder Körperregion eine jeweils einzigartige Zusammensetzung von Mikroorganismen.
Das Mikrobiom des Darms repräsentiert die Gesamtheit aller mikrobiellen Mitbewohner im Darm, vor allem im Dickdarm. In erster Linie sind hier physiologische bzw. schützende Bakterienspezies gemeint.
Schätzungsweise 90% der im menschlichen Körper gefundenen Zellen sind schließlich nicht menschlich, sondern überwiegend prokaryotischen Ursprungs. So enthält der Darm schätzungsweise 100 Billionen Bakterien, etwa zehnmal so viel wie unser Körper Zellen hat, aus einer mikrobieller Gemeinschaft von 1.000 bis 1.500 Bakterienarten.
Eine wichtige Gruppe von Stoffwechselprodukten, die je nach Nahrung von den Darmbakterien ausgeschieden werden und u.a. anti-entzündliche Eigenschaften aufweisen, sind die sogenannten Polyamine. Zu diesen zählt auch das eng mit dem Zellwachstum verbundene Spermidin.
Durch die Verwertung bestimmter Nahrungsmittel, die große Mengen an Polyaminen bzw. dessen Vorläufer-Molekül L-Arginin enthalten, wie beispielsweise Milchprodukte, Gemüse, Fleisch, Fisch und Sojabohnen, sind die Darmbakterien in der Lage, das Polyamin Spermidin zu produzieren. Bacteroides und Fusobakterien, zwei numerisch dominante Bakterienpopulationen synthetisieren im Dickdarm, Spermidin und Putrescin. Bakterielle Polyamine werden im Blinddarm und Dickdarm absorbiert. Pektin, ein lösliches unverdauliches Polysaccharid, regt Darmmikroben an, große Mengen an Polyaminen zu synthetisieren.

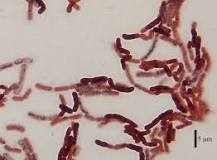
Abb.: BACILLUS MEGATERIUM IST DAS GRÖSSTE BEKANNTE BAKTERIUM UND IST IM DARM DER NACKTMULLE (HETEROCEPHALUS GLABER) FÜR DIE SPERMIDIN-BIOSYNTHESE VERANTWORTLICH. Quelle: https://en.wikipeia.or
Synthese von Vitamin B12
Weder tierische Organismen noch Pflanzen sind in der Lage, Vitamin B12 zu synthetisieren – somit auch nicht der Mensch. Diese Fähigkeit besitzen lediglich Mikroorganismen. Mikroorganismen synthetisieren im Dickdarm des Menschen reichlich B12– und Folsäurevitamine.
LUCA ein biologisches Erfolgsmodell
LUCA – Last universal common ancestor – ist ein biologisches Erfolgsmodell. Wir profitieren noch heute davon. Im Verlauf dieser Evolution-Geschichte traten die entscheidensten Anpassungen (Adaptation) im Stoffwechsel schon zu Beginn des Lebens auf der Erde auf, die es gestatteten durch epigenetische Anpassungen an die Umwelt, die für ein Individuum nötigen Ressourcen aus der Umwelt zu gewinnen.
Die ersten 1,5 Milliarden Jahre nach der Entstehung des Lebens gab es auf der Erde nur Bakterien und Archaeen, und sie sollten das Leben noch mehr als eine weitere Milliarde Jahre dominieren. Bakterien und Archaeen sind beide sehr klein und nur in Ausnahmefällen mit dem bloßen Auge zu erkennen; aber sie kommen in riesigen Zahlen vor und können sich sehr schnell vermehren. In dem riesigen Zeitraum, in dem sie das Leben auf der Erde alleine bestimmten, konnten Bakterien und Archaeen ungezählte Möglichkeiten biochemischer Reaktionen „ausprobieren“ – aus den Lösungen, die die natürliche Selektion überstanden, und die vor allem auf 20 Aminosäuren und aus diesen gebildeten ein- bis zweihundert Eiweißstrukturen beruhen, die die anderen Reaktionen katalysieren, besteht das Leben im wesentlichen noch heute. Spätere Lebensformen sollten die biochemischen Lösungen, die die Bakterien erfunden hatten, kombinieren; aber kaum noch Neues hinzufügen.
Die über 3.5-3.8 Mrd. Jahre antiquierten Stoffwechselwege der S-Adenosyl-L-Methionin- und der Spermidin- Biosynthese, zeigen auch die zwingende Unentbehrsamkeit von S-Adenosyl-L-Methionin und Spermidin als Schlüsselmoleküle des Zellstoffwechsels für den Erhalt der Zell- und Mitochondrien-Gesundheit.
Da die Biosynthese beider Schlüsselmoleküle bereits im ersten Lebensdrittel abnimmt und sie im 80. Lebensjahr zu 80% reduziert ist, ist es ratsam den drohenden S-Adenosyl-L-Methionin- und Spermidin-Mangel, gewissenhaft und frühzeitig durch orale Supplementierung zu ergänzen, um eine gesunde und nicht durch Krankheiten verkürzte Lebensspanne zu sichern.
Ihr
Eduard Rappold
EPIGENETIC BRAIN PROTECTOR – EBP®
enthält 400 mg S-Adenosyl-L-Methionin (Ademethionin)
pro Kapsel (Monatsbedarf pro Packung)


nugenis.eu/shop

SOJAPULVER aus ÖSTERREICH
MIT HOHEM SPERMIDINGEHALT
mehr als 1,7mg Spermidin pro Esslöffel
Inhalt: 40 Portionen
Hergestellt aus gentechnisch nicht veränderten Sojabohnen (NON GVO <0,1%)
abgefüllt in Membrandosen Ø 99×180
geeignet für: Vegetarier, Veganer, Kosher
SOJAPULVER aus ÖSTERREICH
enthält als funktionelles Lebensmittel
SOJA-PROTEIN
essentielle Aminosäuren mit dem
idealen Aminosäure-Score von 100
+
SOJA-SPERMIDIN
erhöht die allgemeine Proteinsynthese um
das 1,5- bis 2,0-fache und
wirkt gegen den altersbedingten Muskelverlust
SOJA-SPERMIDIN induziert die Autophagie und verlängert so die gesunde Lebensdauer der Zellen mit
Anti-Aging-Effekt für alle Zellen
spermidine-soyup.com
www.nugenis.eu/shop
Eduard Rappold
Dr. Eduard Rappold, MSc ist ein erfahrener Forscher und Arzt, der sich seit Jahrzehnten für geriatrische PatientInnen einsetzt. In seinem Bemühen für Alzheimer-Erkrankte eine immer bessere Versorgung zu ermöglichen, wurde er 2003 mit dem Gesundheitspreis der Stadt Wien für das Ernährungszustandsmonitoring von Alzheimer-Kranken ausgezeichnet. Im Zuge seines Masterstudiums der Geriatrie hat er seine Entwicklung des Epigenetic Brain Protector wissenschaftlich fundiert und empirisch überprüft. Im September 2015 gründete er NUGENIS, ein Unternehmen, mit dem er Wissenschaft und Anwendung zusammenbringen möchte. Damit können Menschen unmittelbar von den Ergebnissen der Angewandten Epigenetik für ihre Gesundheit profitieren. Mit dem Epigenetic Brain Protector hat Dr. Eduard Rappold, MSc bereits für internationales Aufsehen gesorgt – auf der international wichtigsten Innovationsmesse, der iENA, wurde er 2015 mit einer Goldmedaille für hervorragende Leistungen zum Schutz vor Neurodegeneration ausgezeichnet. Auf den Webseiten nugenis.eu, epigenetik.at, spermidine-soyup.com und facebook.com/nugenis können Themen zur Epigenetik und Aktuelles nachgelesen werden.